
There is a fundamental structure to the neural system of the animal that is more basic than the neuron. This is the backbone structure that conveys a signal from an originating sensory mechanism to the brain and back to some form of response device. This fundamental structure consists of a series of electrolytic conduits separated at intervals by active electrolytic semiconducting devices designed to process or regenerate the signal.
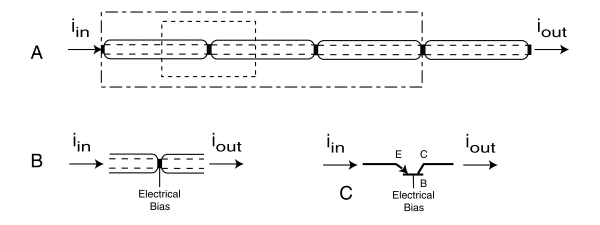
Frame A shows this structure in its simplest form. The labeling hints at an electrical analog related to the signaling function. The dotted line encloses the fundamental building block of the neural system. This block includes a junction, a portion of an electrolytic conduit preceding the junction and a portion of a second electrolytic conduit following the junction. The dash-dot line encloses what could be described as the signaling portion of a generic neuron. A generic neuron frequently contains more than one fundamental building block. Note that the envelope of the generic neuron does not include an integral number of functional units. Frame B points to the fact that there is the possibility of an electrical connection between the surrounding electrolytic matrix and the region defined as a junction as well as to the two ends. Frame C shows the nominal junction replaced by a symbolic element named an Activa, the biological equivalent of a man made transistor.
The Activa is characterized by its three terminals. The terminal connected to the input conduit is known as the emitter (e). The terminal connected to the output conduit is known as the collector (c). The third terminal is known as the base (b).
This basic signaling structure is supported by a metabolic structure, which includes a soma that includes a nucleus and many other sophisticate structures designed to create the original signaling path, maintain that path and also provide other specialized functions indicated by their membership in the broader class of neurosecretory cells. There are a number of simple morphological forms known generically as neurons. Some of these are shown in the following figure.
In the figure, the dark black rectangles between the conduits are all Activas. Some of these Activas are enclosed entirely within one neuron while other occur at junctions between neurons (synapses). In some cases, the internal junctions are known explicitly from the literature, such as the Nodes of Ranvier. Others are only known implicitly from the literature such as the mechanism enclose by the Hillocks of many neurons. In fact, all neurons contain at least one Activa within the body of the neuron. Some specialized cells, such as the photoreceptor cells (not discussed until later) will be shown to contain more than one.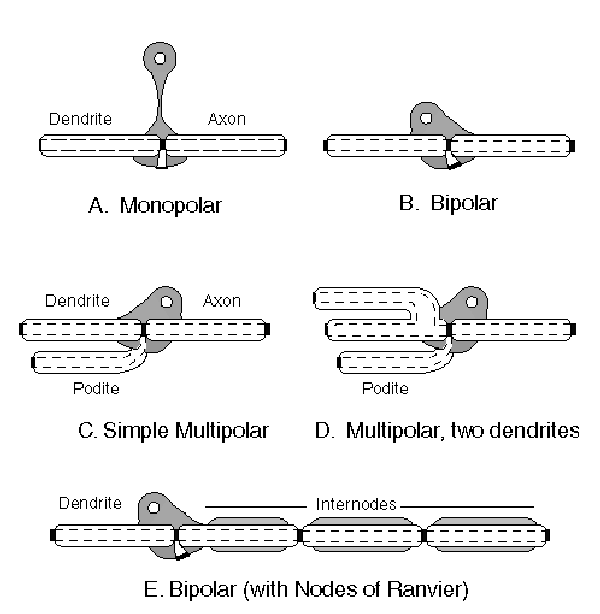
A new element is introduced in these cartoons. The podite (foot in Latin)is a second type of neurite similar to the dendrite. It functions as a conduit connecting the base of an internal Activa with the surrounding electrolytic matrix. The podite has not been recognized explicitly in the literature. However, there are a number of excellent images of the arborization of neurons that clearly show two separate "dendrite like" structures emanating from one soma.
In this figure, some of the podites are shown bent to the left to parallel the dendrite of a neuron. In the completely arborized neuron, the arbors of the podite and dendrite may entangle to a significant degree.
In other configurations, the podite is bent to the right and parallels the axon. In many neurons associated with the signal manipulation function in the retina and elsewhere, these two functional elements are enclosed within a single outer lemma. These neurons are called lateral cells and include the horizontal and amercine cells of the retina.
It is the purpose of this document to provide a framework for understanding the fundamental operation of these different devices by quantifying the physical characteristics of these structures related to signaling and to tabulate the values of the parameters affecting that operation.
Adaptation by animals to satisfy environmental requirements have led to a wide variety of neural forms. Although many authors have worked to identify these forms morphologically, that is not the goal here. Correlation to their morphological form is secondary to their operational characterization. There is one clear dichotomy among the operational forms of neurons, whether they are used to transmit analog or pulse signals over the conduits of the animal. Even this dichotomy is circumscribed by the necessity of at least one type that acts as a transition device interconnecting devices serving these two distinct roles.
There is a second dichotomy found mainly, but not exclusively among those neurons transmitting pulse signals. This is myelination of the output conduit (the axon) to improve its electrical transmission characteristics over extended distances of a few millimeters or more. This characteristic is mandatory because some morphologically defined neurons are known to have lengths measured in meters. In these cases, the so-called axon is in fact subdivided into a series of short conduits separated by active regeneration circuits known as Nodes of Ranvier.
Within the above histological framework, it is possible to characterize all of the functional elements of the animal neural system involved in signaling in considerable detail. The next level of detail concerning the functioning of the neurons appears in the TOPOLOGICAL FORMS OF NEURAL INTERCONNECTION When appropriate, additional specificity can be introduced by referring to this new material in conjunction with that featured in the above figures. To go further requires reference to the main text.